

以前の記事、『一次共生とは?-藻類の起源-』では、シアノバクテリアが真核生物に取り込まれて光合成能を獲得し、その生物が進化と分岐をすることで、スーパーグループ「アーケプラスチダ界」の藻類(灰色藻、紅藻、緑藻)が生まれたことを説明しました。一次共生について詳しく復習したい方は、下記記事をご覧ください。
この記事内で、「アーケプラスチダ界以外の真核藻類は、異なる生い立ちで光合成能を獲得した」とお話ししました。この「異なる生い立ち」というのが「二次共生」です。今回の記事では、この「二次共生」について説明したいと思います。二次共生によって、形態や色、代謝系までもが異なる「藻類の多様性」が生じたのです。
一次共生のおさらい
光合成の仕組みを生み出した原核藻類「シアノバクテリア」を、真核生物が丸ごと細胞内に取り込んで、自分の一部として共生させてしまった現象を「一次共生(primary endosymbiosis)」という事をお話ししました。
ここで取り込まれたシアノバクテリアは、「葉緑体」として細胞内の光合成器官として働いています。一次共生で生まれ、分岐・進化した藻類を「一次植物」と言い、緑藻、紅藻、灰色藻が含まれます。我々が普段目にする植物も、緑藻から分岐しているので一次植物です。
藻類の「葉緑体の共生」を考えるとき、葉緑体の膜の枚数を見てみると、とても面白いことがわかります。シアノバクテリアの細胞膜の枚数は外膜と内膜の2枚です。シアノバクテリアを共生してできた、つまり一次共生でできた葉緑体も、シアノバクテリアを丸ごと取り込んだので、膜の枚数は同じく2枚です。
一次共生の藻類の光合成色素
二次共生をお話しする前に、「藻類の光合成色素」について説明します。
原核生物のシアノバクテリアも、真核生物の藻類も、共通して青みを帯びた緑色のクロロフィルaを持っています。さらに、一次共生で生まれた灰色藻、紅藻、緑藻は、それぞれ特有の光合成色素を進化させました。進化した光合成色素と、クロロフィルaの含有量を変化させることで、藻類の名前に示す色が現れてきたのです。
順に見ていくと、「灰色藻*」は青緑色の藻類です。光合成色素はクロロフィルaと青色のフィコビリンが含まれています。灰色藻の光合成色素は、シアノバクテリアの光合成色素と極めて似ています。
*日本語では「灰色」藻という名称ですが、英語では Glaucophyta(glaucus :地中海の色(sea-green))と訳されます。つまり、英語では「海の緑色」という意味になります。
「紅藻」は赤色の藻類です(一部、青緑色の藻類もあります)。光合成色素は、クロロフィルaと赤色や紫色のフィコビリン、カロテノイドが含まれます。紅藻の葉緑体の色はフィコビリンとカロテノイドが特徴づけています。
「緑藻」は緑色の藻類です。健康食品として人気のクロレラや、ミカヅキモ、ボルボックスなどです。緑藻の葉緑体に含まれる光合成色素には、クロロフィルaとクロロフィルbが含まれています。クロロフィルbは、深い緑色をしています。
二次共生とは
全生物の系統樹では、5つのスーパーグループの「界」と所属不明の生物に分類されていて、藻類は多くの界に点在しています。詳しくは『藻類とは?-実は曖昧な藻類の定義。その理由に迫る-』をご覧ください。

一次共生で生まれた灰色藻、紅藻、緑藻は、上記記事でも紹介したスーパーグループ「アーケプラスチダ」にまとめられる藻類です。アーケプラスチダ界以外の藻類はどうやって葉緑体を獲得したのでしょうか?
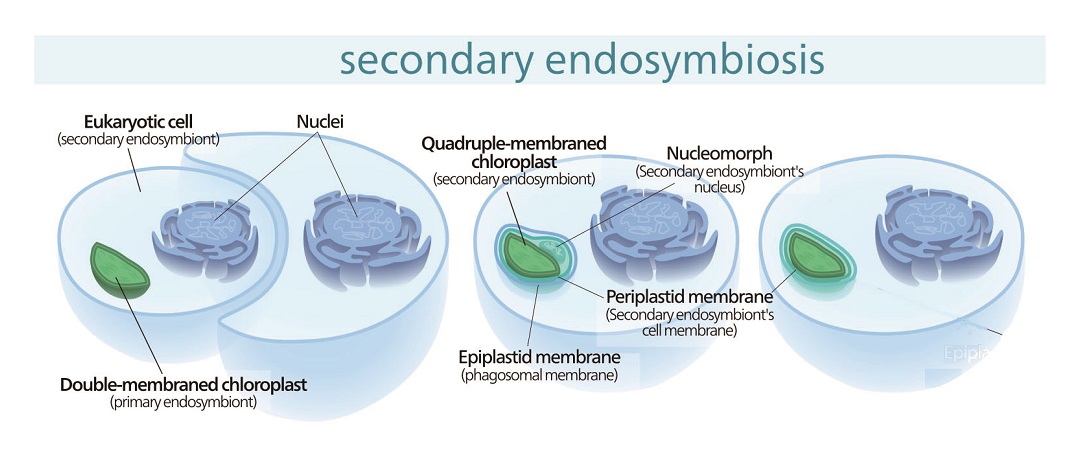
答えは、一次共生で生まれた藻類を、今度はまた別の真核生物が丸ごと細胞内に取り込んで、自分の一部として共生させてしまったのです。この現象を、「二次共生(secondary endosymbiosis)」といいます。二次共生で生まれ、分岐・進化した藻類を「二次植物」と言います。シアノバクテリアと緑藻、紅藻、灰色藻以外の、すべての藻類が二次植物に含まれます。
二次共生で生まれた藻類であることは、どのようにして分かるの?と思われる方も多いでしょう。ここで、葉緑体の膜の枚数と光合成色素を見てみると、なるほど!と驚くのです。
二次共生の成立過程ークロララクニオン藻ー
 (Keeling 2004をもとに作図)
(Keeling 2004をもとに作図)
二次共生について、具体的な藻類を例に説明します。
あまり世間的には知られていませんが、「クロララクニオン藻」という緑色の変わった藻類群がいます。アメーバ状から球状なものまで、形態は様々です。
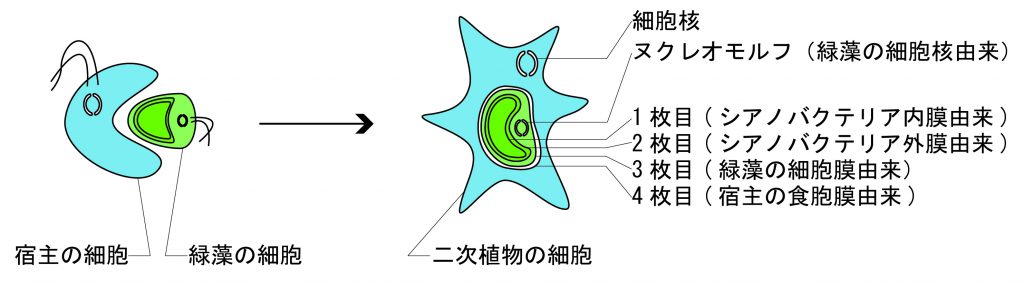
クロララクニオン藻の葉緑体の光合成色素は、クロロフィルaとクロロフィルbを持っているため、一次植物の緑藻と同じであることが分かります。一方で、緑藻の葉緑体の膜が2枚なのに対し、クロララクニオン藻の葉緑体の膜の枚数は4枚あります。クロララクニオン藻の葉緑体には余計な膜が2枚あることになるのです。この余分な2枚の膜の由来が、二次共生を経て獲得された葉緑体である証拠になるのです。さらに、クロララクニオン藻の葉緑体には、細胞核のようなものが含まれているのです。この細胞核のようなものは、葉緑体の内側から2枚目と3枚目の間に存在しています。
光合成色素が緑藻と同じくクロロフィルaとクロロフィルbであること、葉緑体2枚目と3枚目の間に細胞核のようなものが存在すること、そして葉緑体3枚目の膜がそれを包み込んでいること。この葉緑体3枚目の膜が緑藻の細胞膜とすると、クロララクニオン藻の葉緑体の内側から3枚目の膜までに関しては、まさに緑藻と同じ構造になっているのです。つまり、クロララクニオン藻の葉緑体は、緑藻類がそのまま入れ子状態になっているのです。そして残りの葉緑体4枚目の膜は、宿主が取り込むときに包み込んだ食胞膜と考えられています。
このように、膜の枚数から二次共生の成立の過程が垣間見えるのです。
ちなみに、クロララクニオン藻の葉緑体に入っている細胞核のようなものを、ヌクレオモルフnucleomorph(nucleo:核、morph:’形をしたもの)と言います。ヌクレオモルフは元々の緑藻の細胞核に比べると、遺伝子数が減少し、遺伝子密度が高まっていて、はるかに異なっています。ここには、クロララクニオン藻が生きていくうえで、違う生き物である緑藻を葉緑体として制御するために奪えなかった、必須の遺伝子が乗っていると考えられています。
ここでは詳しく述べませんが、一次共生の葉緑体化を完成させた二次植物は、膜の枚数を増やすことでタンパク質の輸送系、代謝系等を発達させてきました。これについては、現在も研究が盛んに行われています。
様々な二次共生
クロララクニオン藻を例に、二次共生の葉緑体の成立過程をお話ししました。他の二次共生によって生まれた藻類も、同様の過程を経て現在の藻類になったとされています。
しかし、取り込んだ一次植物の藻類の制御方法は、各藻類独自に進化させました。制御方法は、どの藻類も試行錯誤したようです。
 (Keeling 2004をもとに作図)
(Keeling 2004をもとに作図)
緑藻を取り込んだ二次植物には、先に話したクロララクニオン藻(リザリア界)とユーグレナ藻(エクスカバータ界)があります。クロララクニオン藻とユーグレナ藻は、それぞれ独立に緑藻を取り込んだとされています。クロララクニオン藻と違い、ユーグレナ藻は葉緑体の膜の枚数は3枚なのと、ヌクレオモルフが無いことが特徴です。ユーグレナ藻は二次共生の過程で、4枚目の一次植物の藻類を捕食するのに使った食胞膜を失ったと予想されます。
紅藻を取り込んだとされる二次植物には、クリプト藻(界の所属不明)、ハプト藻(界の所属不明)、不等毛藻類(ストラメノパイル界。ケイ藻、褐藻、真正点眼藻など複数の藻類が含まれる。)、渦鞭毛藻(アルベオラータ界)があります。紅藻を取り込んだ後、各々の藻類は独自の光合成色素を進化させているため、紅藻と紅藻を取り込んだ藻類の共通性は乏しいです。しかし、二次植物の葉緑体の特徴として、黄緑色のクロロフィルcとカロテノイドを多く含むため、赤色~茶色の葉緑体を持っています。赤色系の光合成色素を多く持つことは、紅藻と紅藻を取り込んだ藻類の特徴といえます。ヌクレオモルフを持つのはクリプト藻だけです。
葉緑体の膜は、クリプト藻、ハプト藻、不等毛藻類は4枚、渦鞭毛藻は3枚所有しています。渦鞭毛藻も、ユーグレナ藻同様に二次共生の過程で、4枚目の食胞膜を失ったと予想されます。クリプト藻、ハプト藻、不等毛藻類は、クロララクニオン藻と同様の二次共生過程を経ていると考えられます。
さらに、この3種の藻類は、細胞核の膜と融合しています。細胞核膜と融合して、4枚目の膜にはタンパク質に翻訳するリボソームが付着しています。リボソームは遺伝情報を持ったmRNAをタンパク質に翻訳にする役割を持っています。葉緑体の最外膜である4枚目の膜が細胞の核膜と融合することで、細胞核に乗っている葉緑体に使われる遺伝子情報を、最短ルートでタンパク質に翻訳して輸送することができるのです。
ちなみに、葉緑体最外膜と細胞核の核膜が融合していないクロララクニオン藻、ユーグレナ藻、渦鞭毛藻による、細胞核に遺伝情報を持つ葉緑体タンパク質の輸送は、各々の藻類で小胞体やゴルジ体を介して複雑な輸送経路を進化させています。
紅藻の二次共生は、クリプト藻、ハプト藻、不等毛藻類、渦鞭毛藻で独立して起こったのか、それとも、1度だけ起こってそこからクリプト藻、ハプト藻、不等毛藻類、渦鞭毛藻に進化していったのか。これは、いまだに解決していない問題です。
上図は後者の、紅藻の取り込みが一度だけ起こったときの二次共生の様子を示したものです。古くから、共生現象は頻繁には起こらないものと思われていて、紅藻系列の二次植物は単系統だと考えられてきました。しかし、紅藻系列の藻類は単系統になる場合、つじつまが合わない部分があるのも事実です。各藻類の仲間には、葉緑体を持たない生物がいるのです。葉緑体を使わない生き方を選択しても、葉緑体の痕跡が残ったり、細胞核に遺伝子が残ったりするものです。
研究が進むにつれて、紅藻の二次共生は、クリプト藻、ハプト藻、不等毛藻類、渦鞭毛藻で独立して起こった、つまり生物界ごとに独立で起こったとする説が主流となっています。
最後に
葉緑体の一次共生も依然不明なところがありますが、二次共生はますます不明な点が多いです。ただ、絶対に言えることは、光合成をする真核生物はみな、シアノバクテリアが作った光合成の仕組みを、一次共生または二次共生によって使っているという事です。
藻類の大半は、一次共生、二次共生によって生まれたのだと思ってよいでしょう。しかし、2度あることは3度ある、と言われるように、「三次共生」、「四次共生」・・・そういった葉緑体獲得様式の藻類も実は存在します。また、別のシアノバクテリアを取り込んだ「もう一つの一次共生」の葉緑体獲得様式をした藻類もあるのです。これらも別の機会に紹介したいと思っています。
「共生」という現象によって、取り込んだ生物の特徴と、取り込まれた生物の特徴が合わさった新しい生物が生まれます。藻類の多様性は、複数の生物界で独立に共生をしたからこそ生まれたのです。逆に、藻類の共生現象が分かると、光合成色素や、タンパク質輸送系など各藻類が独自に進化を重ねて生まれた藻類の多様性の謎が少しずつ解明されてくるのです。
藻類とはなにか?藻類を知れば知るほど、なんなのか分からなくなってきます。
そうやって筆者は、藻類の深みに今もはまっているのです。
参考資料
Keeling, Patrick J. “Diversity and evolutionary history of plastids and their hosts.”American Journal of Botany 91.10 (2004): 1481-1493.
